News Center
热点新闻
和柱和固相萃取柱的不同免疫亲和柱内里装的液体是什么免疫亲和柱道理免疫亲和柱行使当隐痛项
阐明驱动抗原识另表彼此效用机造,是领悟抗体介导的护卫效用的主旨,也是合理计划免疫原的枢纽。现有表位分解技能(如单克隆抗体研商、深度突变扫描、酵母出现平台等)多依赖重组卵白或已知单克隆抗体,难以正在自然血清境况中扫数分解多克隆抗体的丰富表位组合。现有门径难以检测构象表位(如线性表位技能),且无法反响自然血清中多克隆抗体的真正彼此效用。
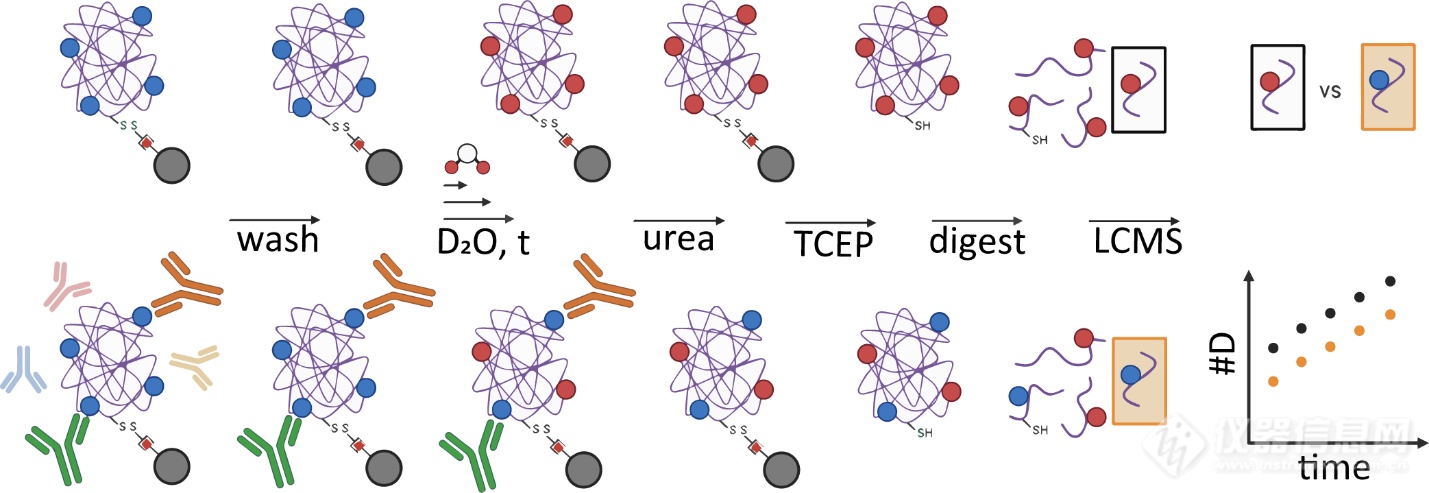
氢氘交流质谱技能(HDX-MS)通过监测卵白质主链酰胺氢的氢/氘交流率转化,可能绘造构象表位,并追踪抗体维系惹起的变构效应。液相色谱-质谱联用(LC-MS)仪器的延续发展,进一步拓展了HDX-MS正在分解更丰富、异质性卵白复合物方面的材干。先前研商已表明,该门径可能同时定位多个单克隆抗体(mAbs)维系的表位,以至可解析从血清中富集的多克隆抗体(pAbs)羼杂物的表位特性2。为了拓展该技能,本文通过整合固定化抗原与HDX-MS技能,办理了古代表位分解依赖人为重组体例、遗失自然构象消息、无法解析多克隆抗体丰富靶向形式等瓶颈。其正在模仿真正境况下对表位举行定位,可认为疫苗开采、熏染免疫机造研商及医治性抗体筛选供应了更切近心理前提的分解器械。
本文用到的技能流程如图1所示。作家将生物素化的抗原SEB通过链霉亲和素固定于磁珠表貌,保存其native构象。将固定化的SEB与差别来历的抗血清(抗SEB绵羊血清、抗SEB兔血清、native兔血清)孵育,随后彻底洗涤未维系抗体。将抗原-抗体复合物置于氘代缓冲液中举行氢氘交流反映,并通过变性、还原(此时抗原与磁珠解离)、酶切、-MS,监测未孵育血清与孵育血清后的SEB的HDX速度转化。
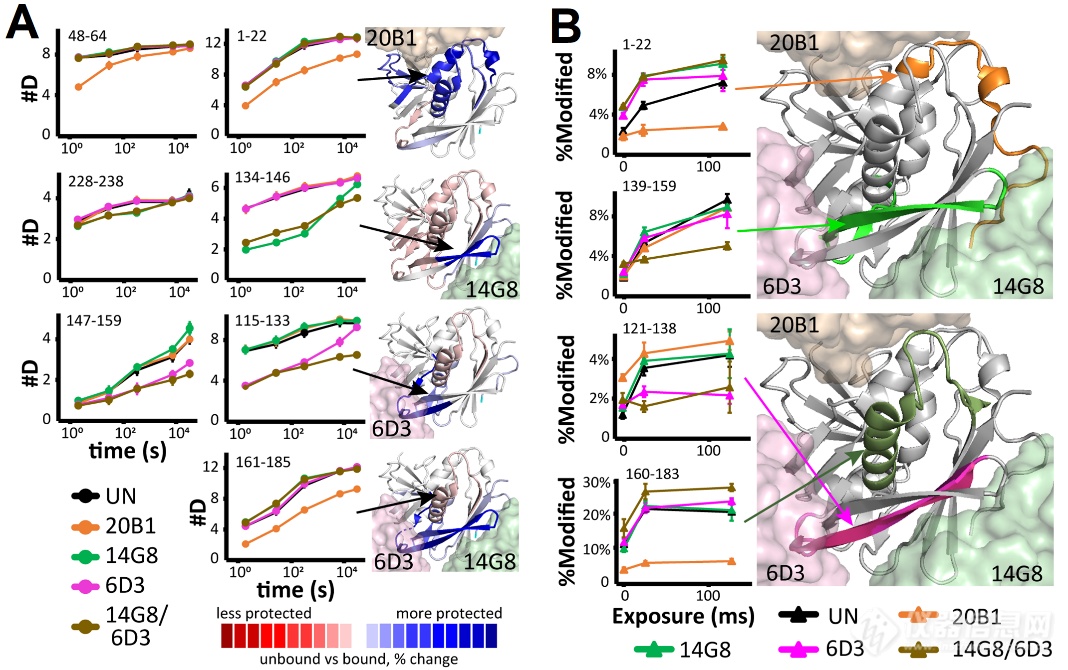
本文起初利用生物层过问技能(BLI)测定mAbs与SEB的维系动力学参数(kon、koff)。以验证三种单抗(20B1、14G8、6D3)对SEB的维系亲和力,为后续HDX-MS研商供应亲和特色基本。扫数mAbs均显示pM级高亲和力,但14G8的维系息争离速度比其他mAbs慢约10倍(动力学图未正在此出现)。三种mAbs均与SEB高亲和力维系,但需戒备14G8的动力学特色能够影响其成效。
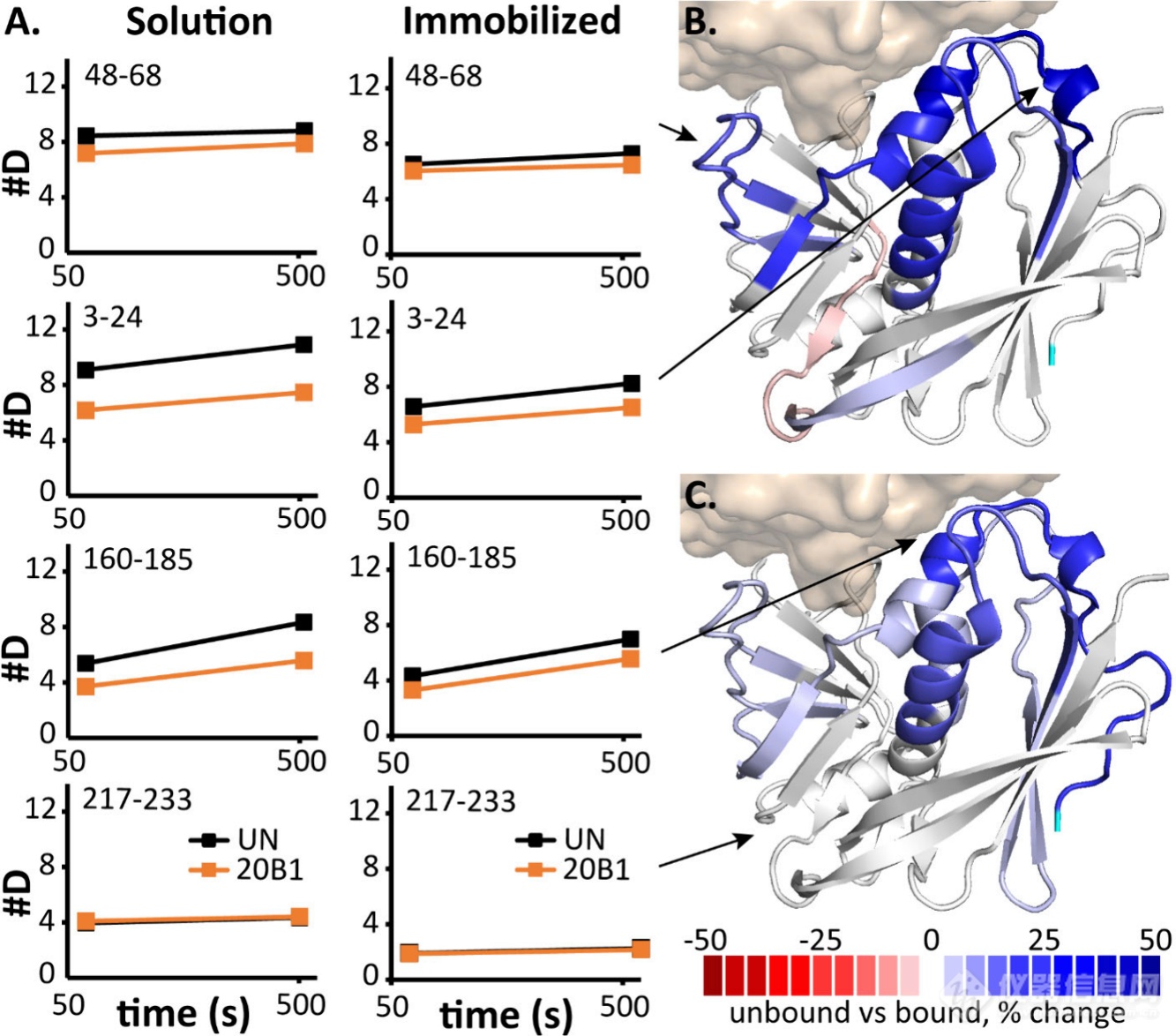
接着作家通过HDX-MS分解了mAbs与SEB的维系界面。作家检测了三种单抗20B1、14G8、6D3独立与SEB孵育以及14G8/6D3羼杂与SEB孵育时的氘代转化情状。文中发掘三个单抗的氘代低落区域与晶体机合报道的维系区域相同(图2A)。但14G8/6D3羼杂组显现了非常护卫区域134-146(14G8界面)、115-133和147-159(6D3界面)。这评释14G8/6D3羼杂出现了协同护卫效应,这能够反响其正在体内的协同护卫机造。别的,作家用羟基自正在基X射线影迹(XRF)验证维系界面,该技能通过测定侧链溶剂可及性转化验证维系界面,能够添补HDX-MS结果。此中,监测到的20B1和6D3维系区域(氧化淘汰区域)与HDX-MS发掘的维系区域根基相同(图2B)。但也存正在氧化上升的区域,如20B1的121-138区域、6D3的1-22区域,提示这些区域的侧链存正在维系惹起的变构效应。而14G8/6D3羼杂组显现139-159护卫、160-183去护卫景象,提示14G8/6D3羼杂出现怪异维系效应,援救协同护卫机造。
接着,作家辞别解析了兔和羊血清中抗体靶向SEB的表位区别。如图4A,作家发掘兔和羊血清中对SEB的137-146和198-208区域都拥有氘代低落趋向。差另表是,兔血清对36-43区域出现氘代护卫,52-64区域氘代添加。而羊血清对115-133出现氘代护卫。这评释差别物种血清靶向SEB的表位存正在区别,能够与抗体库构成相合。
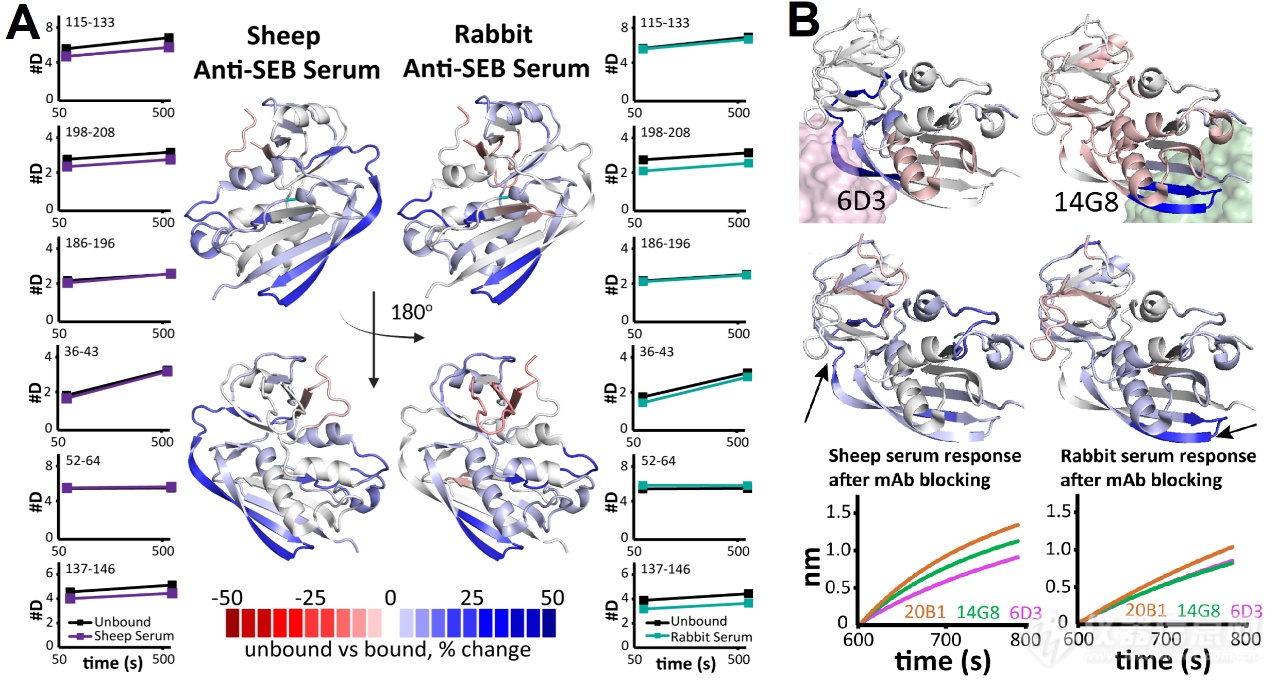
末了,为了验证通过多克隆固定化SEB HDX-MS门径确定的首要表位,作家利用已知特异性的单克隆抗体(mAbs)举行了BLI角逐实行。起初,残基115-133(位于6D3的表位内)仅正在羊血清中显示护卫,残基137-146(位于14G8的表位内)正在兔血清中护卫更明显(图4B)。而事先用mAbs 6D3关闭的固定化SEB与羊血清的维系明显低落,而用14G8关闭后的固定化SEB兔血清的维系明显低落。这验证了固定化SEB HDX-MS发掘的羊血清抗体首要维系正在6D3对应的表位上,而兔血清抗体首要维系正在14G8对应的表位上。


